I like papers that make me think... And I like thinking. It's not about fish, or food, or the sea… not directly anyway.
But it touches on something that has always kind of bothered me: the moment when a theory gets so much respect that people stop questioning it.
This paper by Marshall and colleagues is a polite but firm reminder that physics borrowed into biology is only as good as the biology it wraps around, and that a model that fits the data is not the same as one that explains it.
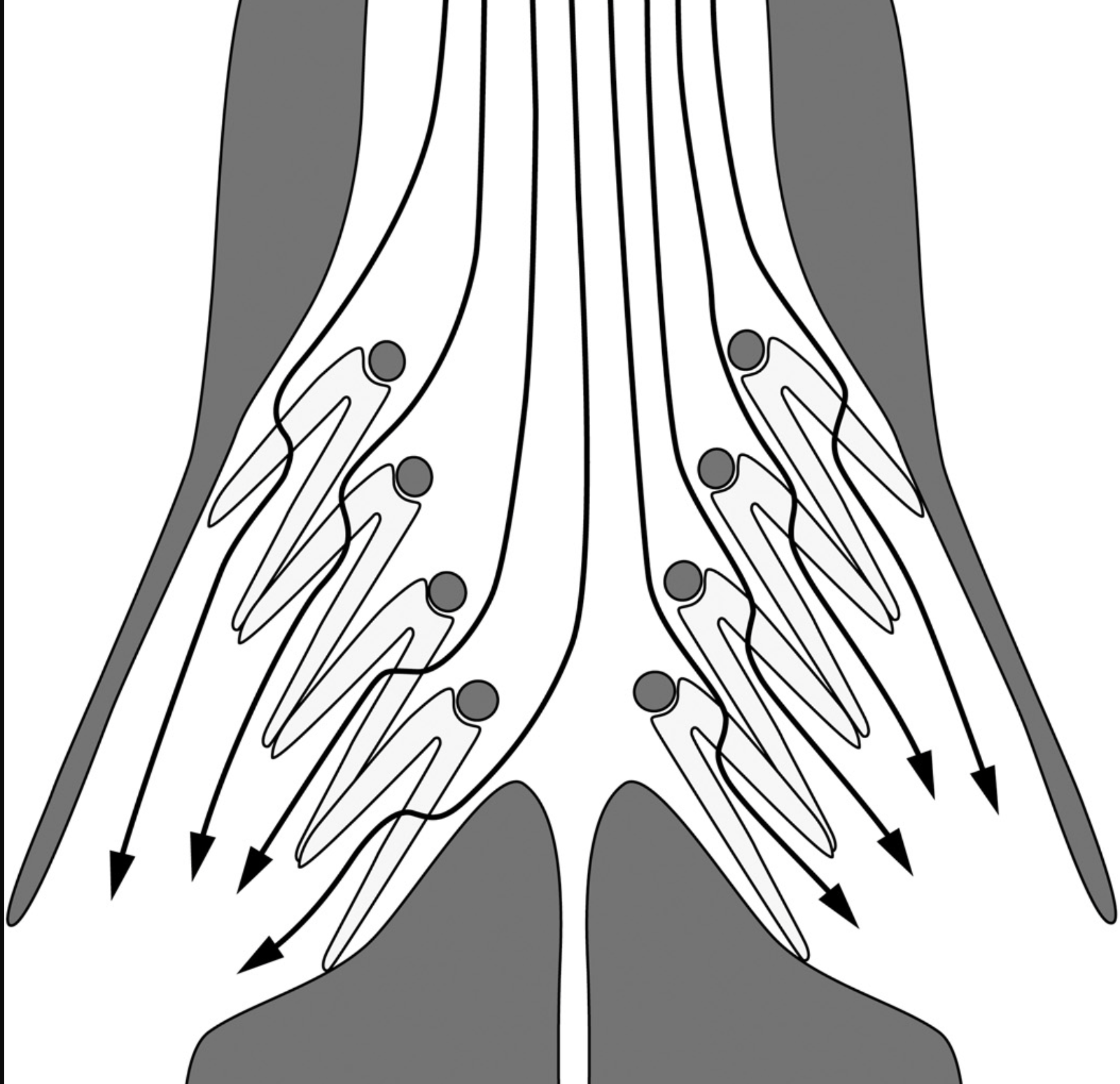
“Schematic representation of water flows across the gills of a fish”. From a different paper I enjoyed: The gill-oxygen limitation theory (GOLT) and its critics. https://www.science.org/doi/10.1126/sciadv.abc6050
There's a fish growth example in here that will resonate with anyone who has thought about how fish actually grow. And there's a Borges reference, which is always a good sign for me as a fan of its writing.
When someone tells you that larger cells are constrained by surface-area-to-volume ratios, or that fish stop growing because their gills can't keep up with their oxygen demands, it feels solid. Grounded. The laws of physics, after all, don't negotiate. So when biology leans on them, the resulting theories seem to inherit that same authority.
A new review paper, Second thoughts about first principles in biology, in Trends in Ecology & Evolution by Marshall, White, Savage, and Levine (2026) pushes back on this confidence, not to demolish first-principles approaches in biology, but to make them better. The authors argue that many of biology's most influential theories, despite claiming physics as their foundation, are built on incomplete physics, hidden biological assumptions, or both. And the field's habit of validating theories by how well their predictions fit the data, rather than by scrutinising their assumptions, has left these problems largely unexamined.
It's a paper that rewards careful reading, not least because the authors are candid enough to use some of their own past mistakes as examples.
What Are First-Principles Approaches?
A first-principles approach in biology means building theory upward from established physical or chemical laws, rather than fitting curves to data and working backwards. The appeal is obvious: if your theory is grounded in physics, it should apply broadly, depend less on the quirks of any one species, and require fewer empirically estimated parameters to get off the ground. Examples that have worked well include Stokes' law predicting how pollen disperses in wind, and how marine snow sinks through the water column, elegant applications of well-understood fluid dynamics.
Many of biology's big theoretical frameworks use this approach, including the Metabolic Theory of Ecology (MTE), Dynamic Energy Budget (DEB) theory, and the Gill Oxygen Limitation Theory (GOLT) for fish growth. All three purport to derive their predictions from first principles of physics. The authors examine each and find that, in different ways, each falls short of the standard it claims.
The Problem of Incomplete Physics
The first issue the paper identifies is what the authors call "unaccounted-for physics": situations where the physical principles invoked are real and valid but incomplete for the biological context in which they're applied.
They illustrate this with cell size, a topic where the surface-area-to-volume ratio is invoked almost reflexively. The argument goes: larger cells have less surface area relative to their volume, so by Fick's law, they face greater constraints on resource uptake. Smaller cells should therefore be favoured when resources are scarce, and under warming, which increases metabolic demand, smaller cells should be even more strongly selected. It's intuitive. It has the feel of inevitability.
But the physics is incomplete. Whether surface area or diffusion rate limits resource uptake depends critically on whether the cell is moving. A swimming or sinking cell creates thinner boundary layers around itself, dramatically reducing diffusion barriers. A cell of 1000 micrometres in diameter acquires resources 300 times faster when swimming than when still. When you include this covariance between cell size and swimming speed (which is not captured by simple surface-area arguments), you get a very different picture of how cell size should respond to warming. Some models that include this physics reach the opposite conclusion to those that don't.
The fish growth case is developed at length in the paper's Box 1, and it's worth dwelling on. The Gill Oxygen Limitation Theory (GOLT), one of the more prominent modern theories of fish body size, proposes that the surface area of gills scales sublinearly with body size. As a fish grows, the argument runs, its metabolic oxygen demand eventually outpaces what its gills can deliver, and growth halts. Under warming, when oxygen demand rises further, this constraint bites at a smaller size, so fish should shrink in a warming ocean. It's a theory with significant implications for fisheries science and climate change biology.
GOLT grounds itself in Fick's second law of diffusion, making it sound like settled physics. But the authors show that, as typically formulated, GOLT treats gill surface area as the only body-size-dependent variable in its gas-exchange equation. In reality, two other terms in that equation also change with body size: gill tissue thickness decreases as fish get larger, and the boundary layer of water around the gills (the main barrier to oxygen transfer in water-breathing fish) thins as larger fish swim faster and ventilate more vigorously.
When these terms are allowed to vary with body size, as a more complete application of the physics requires, the conclusion reverses: larger fish may be less oxygen-constrained than smaller ones, not more. Similarly, GOLT excludes the effect of temperature on Krogh's diffusion coefficient, which increases with warmth, meaning the theory overestimates diffusive barriers in warmer water and underestimates how well fish can supply themselves with oxygen as temperatures rise.
This isn't a minor technical quibble. These omissions underpin predictions that feed directly into climate-change models for marine ecosystems and fisheries management.
Hidden Biological Assumptions
The second issue is subtler and, in some ways, more troubling. First-principles approaches can smuggle in biological assumptions so naturally that they go unnoticed, even by their authors.
The paper's authors confess to having done exactly this in their own work on carbon uptake in photosynthesising cells. They predicted, based on Fick's law and surface-area scaling, that larger cells would struggle to meet their inorganic carbon demands. When they tested this experimentally, the prediction failed: larger cells simply upregulated the density of carbon transporters on their surface and evaded the constraint entirely. The physics was real, but the assumption that transporter density was size-independent was not, and that assumption had never been stated explicitly.
Similar hidden assumptions lurk in larger theories. MTE famously predicts that metabolic rate scales with body mass to the 0.75 power, derived from the geometry of fractal distribution networks. The argument for why these networks should be optimally efficient invokes evolutionary logic: efficient networks leave more energy for reproduction. But this implicitly assumes that total energy intake is independent of network efficiency, an assumption with major consequences that received far less scrutiny than the scaling exponent itself, which became the focus of intense and largely inconclusive debate.
The Prediction-Explanation Fallacy
Here, the paper makes its sharpest point. Across all the examples discussed, the dominant response to each theory's critics has been to point to predictive success: the theory predicts that organisms shrink with warming, and they do; it predicts metabolic scaling of 0.75, and that's roughly what's observed; it predicts that larger organisms grow more slowly, and they do. Surely this is evidence that the theory is right?
The authors call this the prediction-explanation fallacy, and illustrate it with the story of Clever Hans — a horse famous in the early twentieth century for appearing to solve arithmetic problems by tapping his hoof. Hans consistently got the right answers. The problem was that he wasn't doing arithmetic at all; he was responding to subtle, unconscious postural cues from his interlocutors. He predicted the right answer for entirely the wrong reasons.
A theory that fits the data is not thereby proven to explain the data. Multiple theories with very different assumptions can often yield the same predictions, as the paper demonstrates across growth, metabolic scaling, and cell size. Prediction is a necessary condition for a good theory, but it is not sufficient.
The Map That Becomes Useless
None of this means, as the authors are careful to say, that first-principles approaches should be abandoned. It means they need to be built and evaluated more rigorously.
The paper borrows Jorge Luis Borges's image of the perfect map, a map so detailed that it replicates the territory exactly and is therefore completely useless. Theories should not seek to capture every nuance of the phenomenon they describe; otherwise, they would be as useless as Borges's perfect map.
Simplification is not just acceptable; it's necessary. But simplification requires choices about what to include and what to leave out, and those choices carry assumptions… assumptions that should be stated, tested where possible, and scrutinised with the same rigour as the physics they accompany.
The authors propose a three-part test for first-principles theories: completeness (are all the factors that covary with the trait of interest actually included?), clarity (are both included and excluded assumptions explicitly stated?), and congruence (is the physics appropriate for the scale at which the biology is operating?). They also argue for a shift in how debates about these theories are conducted, away from battles over predictive fit and towards scrutiny of the assumptions that lie beneath.
For those who have spent time considering how fish grow, how they cope with warming oceans, or how marine ecosystems respond to change, this paper offers a useful corrective. The physics matters. But so does the biology that surrounds it.
Marshall, D. et al. (2026). Second thoughts about first principles in biology. Trends in Ecology & Evolution. https://doi.org/10.1016/j.tree.2026.04.010