Back in the late 80s when I worked as an observer in the commercial fleet and then a science technician in fisheries research vessels, I did a lot of sampling. The usual job was measuring, sexing and extracting the otholites for later analysis, but then one of the subsamples was what we called “Trophic Ecology” (The study of the feeding relationships of organisms in communities and ecosystems - from the greek τροφὸς: to nurse/feed). The whole idea of this type of work is to understand who eats who in the ecosystem, and how a potential change in abundance at any trophic level could impact the rest.
So after butchering the fish to sex it and removing the otolithes from the cranium, we will get right into the guts, and analyse the stomach content for identification and weight of what ever was recognisable… (we got trained into identifying non digested features).
Not the most charming job in fisheries biology, and particularly with bad weather. But the results where quite critical for the wider job and understanding of the “whole picture”.
So I was immediately drawn to a report published by SPC a few weeks ago based on results of of around 12000 stomach analysed over the last decade. I'll transcribe below some of its methodologies and conclusions.
Building the warm pool ecosystem model
SPC analysed the over 12,000 predator stomachs collected and analysed since the monitoring began. Results of these analyses have been incorporated into a trophic model that describes the warm pool ecosystem and allows forecasting the dynamic responses of the ecosystem to simulated changes in fishing effort through time (Ecopath with Ecosim).
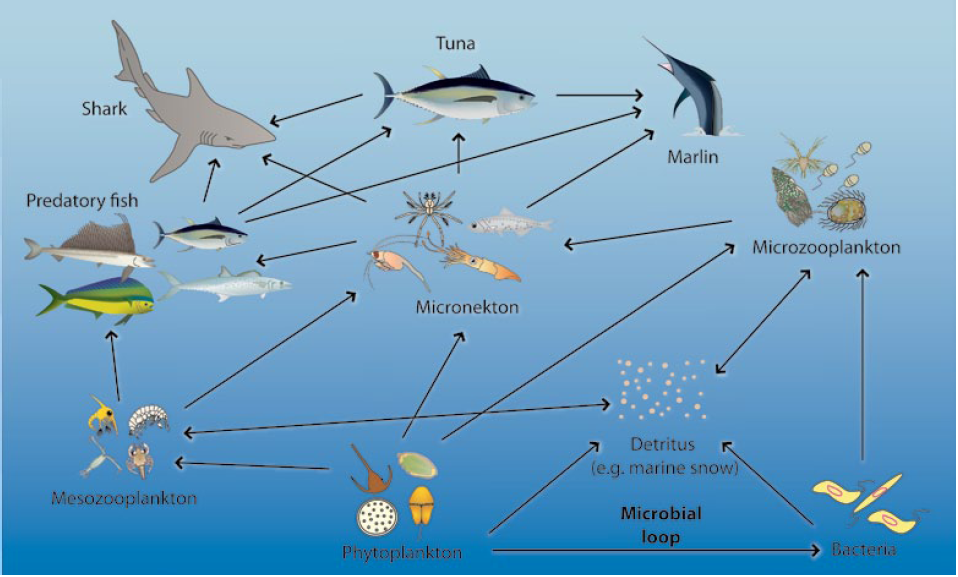
Simplified view of the generalised food web supporting tuna and other large pelagic fish in the warm pool. Note that, at the bottom of the food web, both phytoplankton (microscopic plants) and ‘marine snow’ (phytoplankton and zooplankton remains decomposed by bacteria, also known as detritus) contribute trophic inputs.
The modelled simplified ecosystem was composed of 44 groups: fisheries discards (1 group), detritus (1), phytoplankton (2), zooplankton (2), forage/prey groups (epipelagic, mesopelagic, bathypelagic, migrating or not, fish, mollusc, crustaceans) (11), bycatch species (rainbow runner, pomfret, opah, lancetfish, escolar and oilfish, small tunas, dolphinfish, wahoo) (8), tuna (albacore, skipjack, yellowfin, bigeye of different size classes) (8), sharks (oceanic white-tip, silky, blue, mako and other sharks) (5), billfish (swordfish of two size classes, striped marlin, blue marlin and other billfishes) (5) and turtle (1).
Four fisheries (longline, pole-and-line, purse-seine unassociated schools, purse seine associated schools) were included in the model.
Nine scenarios of fishing effort were explored. They comprised measures designed to reduce/increase the catch of the bycatch community and measures designed to reduce/increase the harvest of tuna by (a) altering the amount of longline fishing and purse-seine fishing, both unassociated (free schools) and associated with fish aggregating devices (FADs), and (b) by simulating the implementation of bycatch mitigation measures. Results were projected for 2026 and 2046.
How does the warm pool ecosystem work and what are its key dynamics?
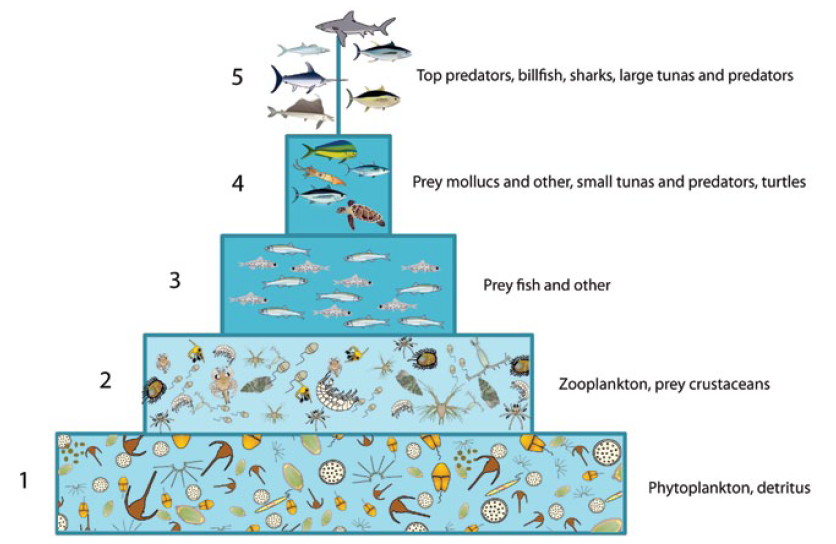
The warm pool ecosystem model is characterised by a large number of trophic links between groups and a diverse pool of prey on which a wide diversity of predators is feeding.
The majority (74%) of the ecosystem’s biomass is in phytoplankton and zooplankton (trophic levels TL 1 and 2), whereas the industrial fish catch (tuna and bycatch) are in TL 4 and 5 at the top of the food web, representing less than 8% of the total biomass of the pelagic ecosystem.
The most important keystone group in the warm pool ecosystem model is small yellowfin tuna, due to its high production and consumption values and its diverse diet. The next most important keystone groups are the prey organisms, which have high production values as predators, but are also important prey for a range of larger fish such as tuna and marlin.
Potential impacts of fishing strategies on the whole ecosystem
The modelled ecosystem was resistant to considerable disturbance from fishing. They suggest that this is related to the considerable diversity of predators in the food web that consume a wide range of prey. Maintaining the diversity contributes importantly to the sustainability of the system.
The structure of the ecosystem was most sensitive to changes in the biomass of prey groups (e.g. small pelagic fish, such as anchovy) because these mid-trophic level species are important prey for tuna, as well as being predators for lower trophic levels, such as zooplankton. Hence, variations in prey availability and quality in relation to changes in the climatic conditions will affect the whole ecosystem and the fisheries.
The simulations showed that groups comprising longlived, bycatch species with low productivity, such as sharks, opah and billfish, are most likely to be affected by changes in purse-seine and longline fishing effort.
Increases in purse-seine fishing on FADs results in greater mortality of sharks and decreases in the biomass of some species and size classes of tuna. This scenario had the most negative impact on the ecosystem. Conversely, reductions in purse-seine fishing on FADs increases the numbers of sharks, although such benefits are not as pronounced when purse-seine fishing effort on FADs is transferred to purse-seine fishing on free schools of tuna.
Increases in longline fishing result in greater mortality of sharks, opah and some billfish species. The negative impact on opah and billfish is also observed when longline fishing effort is unchanged but shark mortality is decreased by the implementation of shark mitigation measures. The simulations to date suggest that some species of the ecosystem will benefit from variations in fishing effort and others will lose; managers will have to define which groups of species are expected to benefit.
It is also apparent that no single indicator is able to provide a good representation of the responses of the ecosystem to changes in harvest. This reflects the complexity of the ecosystem. The use of a variety of indicators is likely to be required to detect the full range of impacts from alterations to harvest strategies.
As with tuna stock assessment models, use of the best available data is critical. Continued and expanded monitoring of catch and discards for bycatch species by observers (at sea or electronic) is critical for further model development and improvement. Similarly, expanding fisheries monitoring programmes to include prey species through predator stomach collection as a routine observer duty is necessary to spatially disaggregate the model.
Further reading
Allain V., Griffiths S., Bell J. and Nicol S. 2015. Monitoring the pelagic ecosystem effects of different levels of fishing effort on the western Pacific Ocean warm pool. Issue-specific national report. Oceanic Fisheries Programme, Secretariat of the Pacific Community, Nouméa, New Caledonia.
http://www.spc.int/oceanfish/en/publications/doc_details/1376-monitoring-the-pelagic-ecosystem-effects-final